


We didn’t realize it at the time, but the first nail we hammered into the lid on the coffin of our scientific career was a paper that we wrote about using cladistic phylogenetic analysis to study the Devonian fossil Archaeopteris.
Abstract
Archaeopteris is now known as a whole plant (Beck, 1960) that is characterized by having gymnospermous secondary wood (pycnoxylic) and pteridophytic (free sporing) reproduction. These characteristics also serve to delimit the progymnospermopsida, a lineage arising from Trimerophyte ancestors before the mid-Devonian, and extending Into the early Carboniferous. The progymnosperms as a whole are regarded as the ancestors of all gymnosperms, the Aneurophytales possibly giving rise to the Pteridospermales and Glossopteridales, and the Archaeopteridales to the Coniferales.
Considerable variation exists within the genus Archaeopteris, in terms of both reproductive and sterile appendage morphology, which has caused Beck (1961) to suggest that the variation may represent more than one genus, and possibly several. However, no estimate is currently available that adequately assesses the interspecific, and intraspecific variation, not to mention the variation that may exist within a single individual.
Introduction
Archaeopteris was initially described in 1871 by Sir J. W. Dawson, along with other genera (Cyclopteris) that have subsequently been synonymized with Archaeopteris (Arnold, 1939). In order to save space, the reader is referred to Gensel and Barnett-Lawrence (in press) for a detailed summary of the history of the genus.
In the first half of the twentieth century there were as many a dozen species of Archaeopteris reported when in 1941 Krausel and Weyland published a review of the genus. Their findings reduced the number of species to between 4 and 6, that fit currently known variation patterns well. The UNC Archaeopteris collection contains specimens that span all of Krausel and Weyland’s (1941) concepts, and also contains additional information.
The function of this study was to phylogenetically analyse the existing variation in Archaeopteris (Dawson), using as OTUs the species concepts of Krausel and Weyland (1941), in the expectation that this would reveal many shortcomings and problems with this approach that could be improved upon and dealt with during the longer time span of a Doctoral thesis.
Considerations
Why phylogenetic analysis? What evidence is there to suggest that any relationships actually exist between these highly fragmentary specimens. How do we know that all of the variation we see couldn’t be found within a single live individual, or that the specimens we have are actually a sampling from different genera, families or even orders of mid-Devonian plants. Of course we don’t know, therefore phylogenetic analysis is the most obvious way to find out what this variation means. It may be that in “real” terms, 50% of our specimens are of a single species, and the other 50% are sampled from across other genera, but in relative terms, 50% of the specimens are more related to themselves than they are to the other 50% of the specimens. The absolute “distance” between any of these individuals is irrelevant on a relative scale.
Classifications derived from phylogenetic analysis require a grouping and a ranking component (Mishler and Brandon 1987), and therefore, a phylogenetic analysis carried out using “perfect” data should be expected to reveal that in the above case, 50% of the taxa form a (probably highly unresolved) monophyletic group, and is sister to the other 50% of the specimens which have other relationships. If this data is also supplemented with an analysis of variance, and possibly other multivariate analysis (such as principal component analysis and cluster analysis), using statistically significant population sizes, It may be possible to discover that the monophyletic group containing 50% of all specimens constitutes a species and not a genus or family.
The problem then, is how to get the most perfect data possible.
Methods used In the current study.
Part 1 OTUs (Operational Taxonomic units)
As mentioned above, the operational taxonomic units (OTUs) in this study are the species concepts of Krausel and Weyland (1941) with some modifications where appropriate or additional information is available.
The OTUs as used are as follows:
Table 1.
- A. Leaf margin entire, at most crenulate, slightly sinuous, sometimes cut but even then the sides are entire.
- Leaves well separated from each other, wedge to more or less fan-shaped, lateral margin more or less straight, often longer than 2 cm (max 7 cm). A. obtusa
- Leaves more closely spaced, somewhat overlapping, distal margin more or less rounded, often longer than 2 cm. A. hibernica (A. latifolia)
- Leaves closely spaced, covering each other by as much as 50%, inner side margin often crenulate, longer than broad, smaller than 2 cm. A. halliana (A. jacksoni, A. rogersi, A. gaspiensis, A. minor also A. roemeriana Kendrick and Fairon-Demaret, 1991)
- B. Leaves more or less incised, also split up on the lateral margins.
- General outline of the leaves is rounded at the top, incised, the Incisions as deep as to the middle of the leaf, lower part of leaf is not split, often less than 2 cm. A. macilenta (A. sphenophyllifolia ::: >2 cm)
- General outline of the leaf rounded at the top, the incisions cut to the base of the leaf. Could be considered as the early stages of webbing of the leaf. A. fissilis
Modified from Krausel and Weyland 1941
Part 2 Outgroup
Choice of the outgroup for this study was luckily obvious. Within the group progymnosperms the Archaeopteridales is accompanied by the Aneurophytales and the Protopityales (Stewart, 1983). Of the two the Aneurophytales is the outgroup of choice because it possesses characters that under some concepts may be considered primitive to those of the Archaeopteridales and, though not reliable due to obvious problems with sampling error, is found to occur earlier in the Devonian than any Archaeopteris species.
Part 3 Characters
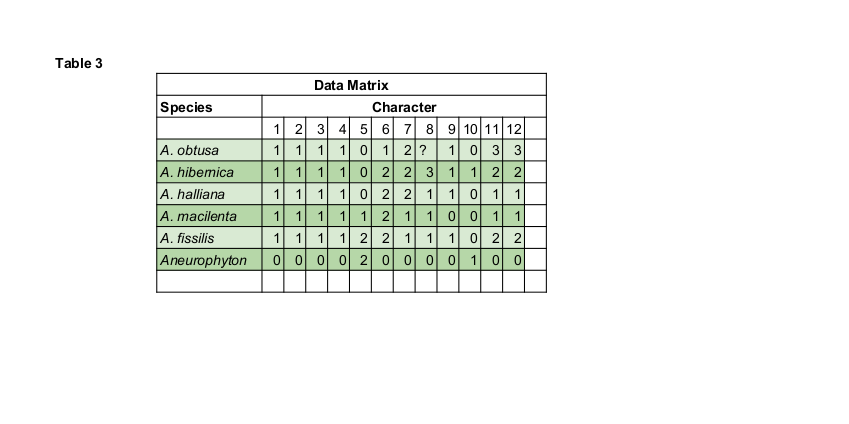
- Stele type. 0; actinostele (Aneurophyton), 1; eustele (Archaeopteris) It may be justifiable to order this with 0 = primitive and 1 = derived.
- Homospory/heterospory. 0; homospory (Aneurophyton), 1; heterospory (most if not all Archaeopteris), may be ordered.
- Terete or planated branch unit. One of the characters that it may be justifiable to order based on the telome concept of Zimmerman (1952). According to this concept, initially undifferentiated, ultimate appendages are spirally arranged upon second order branches that are spirally arranged upon a tertiary branch and so on. As selective pressure for higher efficiency photosynthesis takes place the 3-D branch unit (telome truss) becomes flattened (planated) to better position the photosynthetic apparatus.
- Once planation has begun or has taken place, webs of photosynthetic tissue appear between ultimate appendages forming eventually a lamellar structure (leaf). Aneurophyton and the Archaeopteris species show a similar trend, from Aneurophyton showing the primitive conditions of terete branch unit, and undifferentiated 3-D ultimate appendage arrangement. All of the Archaeopteris species show at least planation of the arrangement of leaves upon the penultimate appendage, and the leaves themselves are planated. Even in A. fissilis there is some degree of webbing between the “veins” of the leaf. A. halliana and associates possess the derived fully formed leaf.
- Incision depth. This character was included as the alternative hypothesis regarding ordering of the leaf margin data according to the telome concept of Zimmerman (1952) as outlined above. 0; entire margin, 1; incisions to 50% leaf length, 2; incisions to leaf base.
- Leaf shape: 0; no leaves, 1; wedge-shaped, 2; elliptical-obovate
- Margin features: 0; no margins, 1; margins only, 2; margins revolute. This is one of those characters that really has been included for the sake of including it.
- Sterile tops. The fertile branch units are in many species of Archaeopteris associated with distal and interspersed vegetative appendages (leaves) that are in the same ontogenetic spiral as the fertile appendages. Krausel and Weyland (1941) distinguish between several slightly different forms of these leaves. O; not present, 1; simple, 2; simple or forked, 3; simple to many times divided. Again, this character has been included purely because it has been reported. Considering the sample sizes available to Krausel and Weyland, and the type of interspecific and intraspecific variation that are known to exist, it seems to me unlikely that this actually represents a character. At the time of Krausel and Wayland’s (1941) work there were no reports of fertile remains of A. obtusa. Such remains have now been discovered and reported (Gensel and Barnett-Lawrence, in press) although dégaging of the distal portion of the specimen by the current author became so difficult, and the remains so hard to trace through the rock, that it is not known whether or not sterile foliage was present at the tip.
- Sporangia number. This refers to the number of fusiform sporangia per fertile unit. There is considerable variation and overlap between the specimens.
Table 2.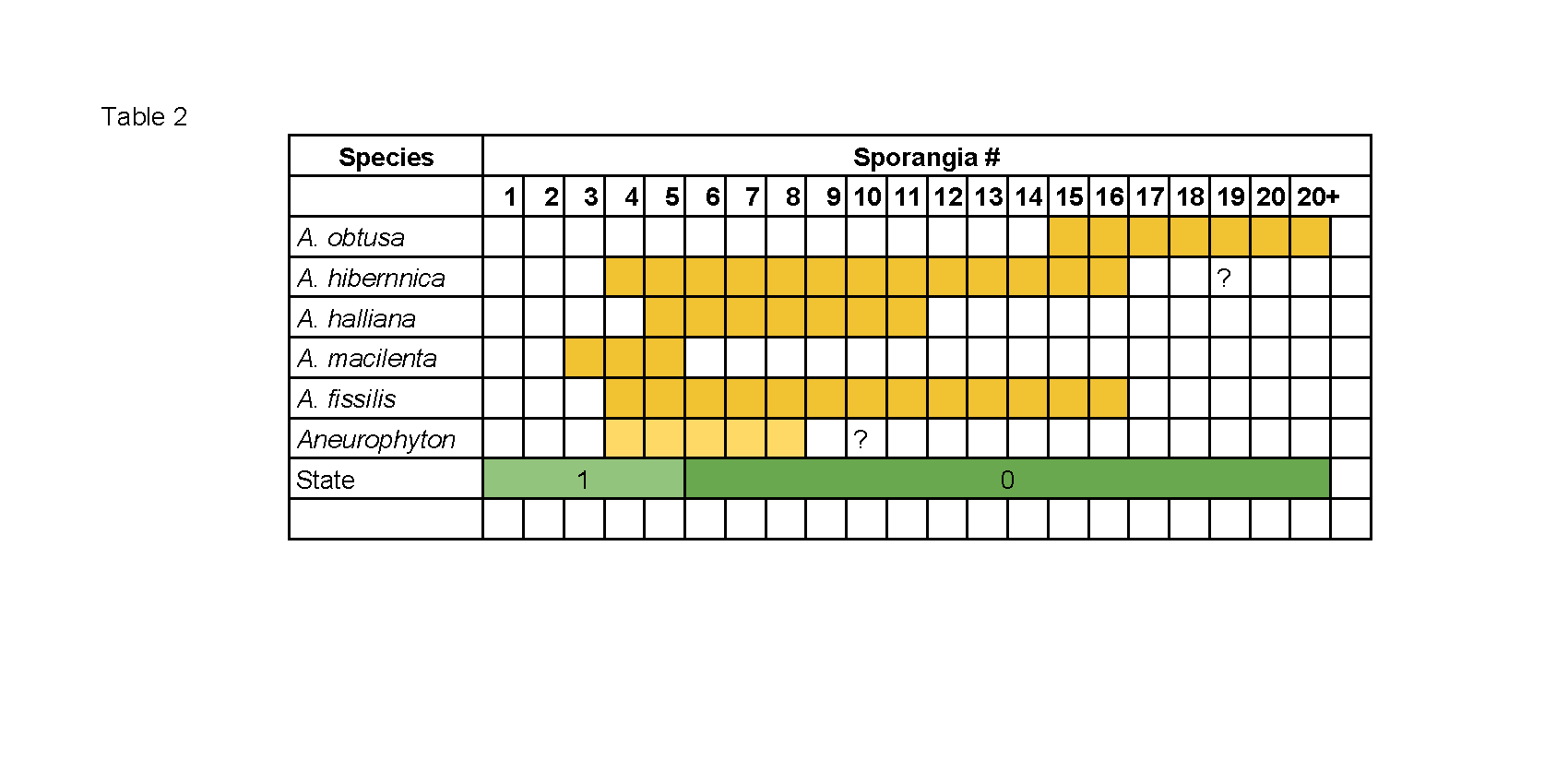 Taking the only clear break in this variation and using it as a character is once again on the shaky side of things, but there is so little to work with that we may just as well.
Taking the only clear break in this variation and using it as a character is once again on the shaky side of things, but there is so little to work with that we may just as well. - Sessile or short stalked. This is in reference to the individual fusiform sporangia and their positioning upon the fertile unit. This is another character of such dubious nature that it is difficult to judge its value. As with other characters indicated as being possibly either uninformative or figments of the human imagination it is included because this Is all that there is.
- Leaf spacing, and Leaf size. Although there are severe weaknesses with regards the use of size criterion in phylogenetic analysis, size based characters were supplied for use if desired due to their critical role in the species concept delimitation of Krausel and Weyland (1941 ).
- It was thought that they may be of use in helping to resolve the polycotomys particularly within the A. halliana clade.
Phylogenetic Analysis
It only takes a glance at the list of characters and the data matrix to instantly realize that there is almost nothing to go on. The extreme paucity of character data leads the tree to be so dominated by certain characters that it in essence becomes just a character state tree. It was found that using both Lundberg rooting and outgroup rooting, and by altering the characters included to test both of the hypotheses of leaf structure development, and using different weights of the size criterion, that the strict consensus trees in every case were very unresolved. There is a monophyletic group containing the A. halliana complex (unresolved). Which is sister to unresolved positions of the A. macilenta types. Archaeopteris is sister to Aneurophyton. The trees from two runs of PAUP are included.
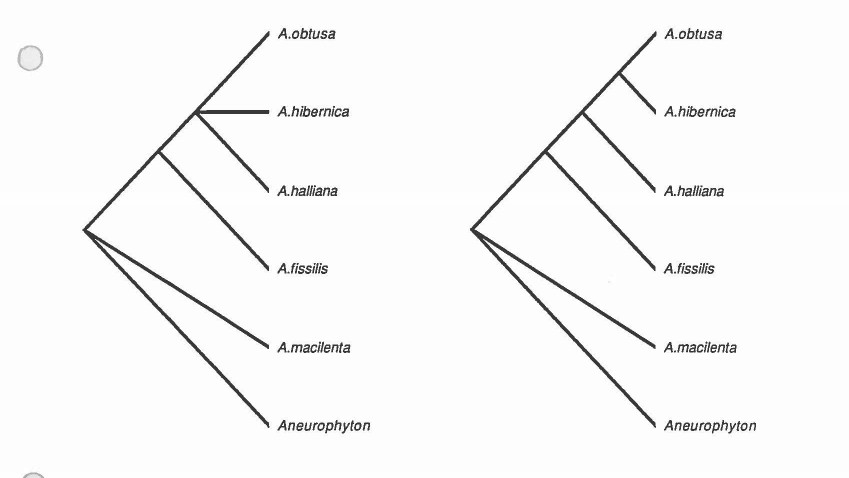
PAUP run 1
Exclude Characters: 5, 11 & 12
Outgroup: Aneurophyton
Ordered characters: 1-4, 6-10
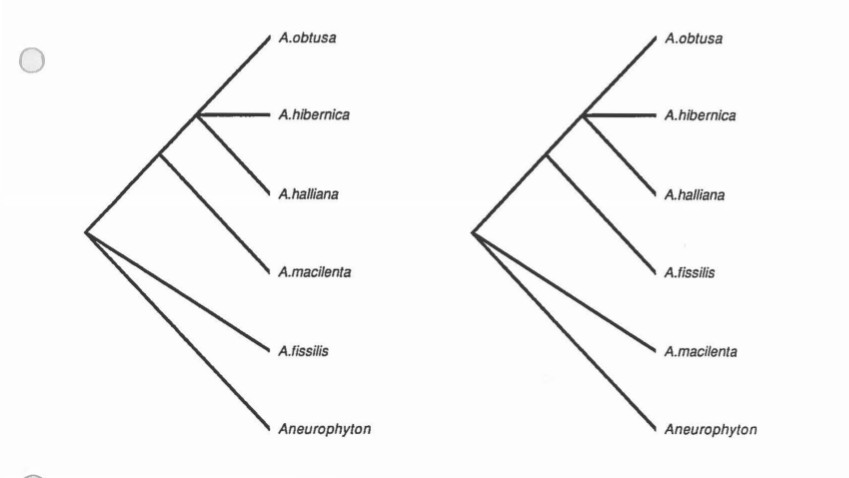
Paup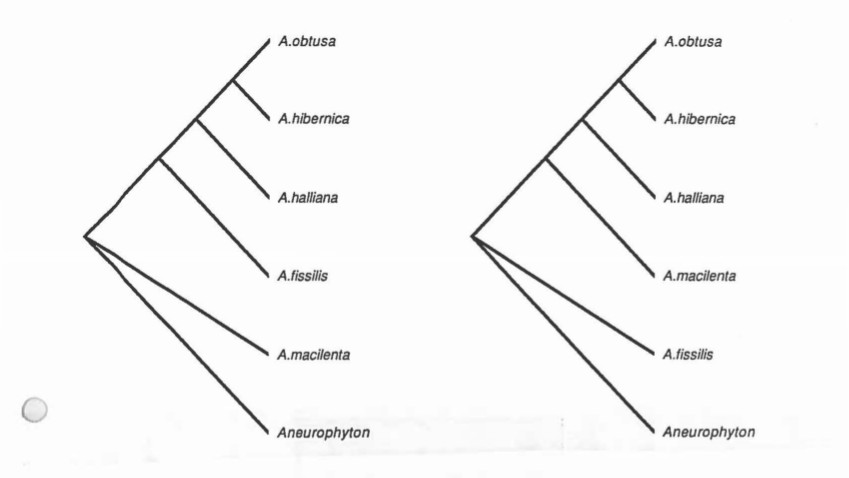
Paup run 2
Exclude characters: 6, 7, 11 & 12
Outgroup: Aneurophyton
Ordered characters; 1-5, 8-10
Discussion
What modifications can be made to actually get some real information out of this study? Firstly, regarding the OTUs. Because of the nature of the specimens from which data was collected, there is no frame of reference as to the ranking of any variation that is seen. The entire variation of Archaeopteris could represent merely the change in leaf morphology from sun to shade leaf. Alternatively it may represent a very diverse, distantly related, polyphyletic (very likely) group. This in turn means that representing this variation with a series of species concepts is highly misleading. In this type of analysis, species concepts should be allowed to generate themselves by providing the software with enough data to be able to distinguish between naturally occurring (monophyletic) groups. How could this be achieved? In order to carry out a perfect phylogenetic analysis of a lineage it would be ideal if it were possible to input every single individual as an OTU. Assuming that sufficient data on synapomorphies was available then at least the higher ranks of the resulting cladogram would be expected to be fairly accurate. In order to provide a sufficiently large sample size it is considered that inputting individual specimens as OTUs, and having no preconceived Ideas about species concepts is much more likely to divulge information about real variation patterns.
Unfortunately however it has been seen that just as much a barrier to phylogenetic analysis is the poignant lack of character data. Not only are there far too few characters, the ones that are available are so cut and dried that they provide almost no information about anything except the fact that a certain concept was used to order them. What can be done about this problem? Technology to the rescue! It is considered that the relatively recently developed methods in morphometrics will be able to provide the character data that Is so needed. It is felt that simply coding leaves as to whether or not leaves have entire or incised margins falls far short of the real situation. There is obviously a fairly complete transition between the two most extreme forms of leaf type, A. obtusa and A. macilenta, that if described in very minute detail could provide the researcher with character sets that approach the size of those being used in molecular phylogenies. Imagine a data matrix with 500+ specimens as the OTUs and 1000+ binary characters that provide maps and plans of the morphologies of vegetative and fertile structures.
In part this information could be provided by very closely analysing the venation pattern that is often preserved with these specimens. For example, it is assumed that the fine hair like processes that compose the leaf margin of the A. macilenta group are vascularized, but the position of the vein tips in A. halliana is not known, are the vein tips associated with the clefts or lobes of the crenulate margin. By using as many homologous points on the leaf (vein branches, process tips, etc.) as possible as a framework, very detailed maps could be produced, the points of which could be coded as synapomorphies.
The modular growth of plants may be of use in the plotting of movements of homologous structures. If for instance a branch unit was in putted into the data matrix as a map of its modular units (starting from ultimate appendages and working proximally), then it would be comparable to any other branch unit on that basis.
Microfossil evidence may be available to supplement this data. Cuticle and spores have very great potential to be preserved, and if a sufficiently large-scale study was undertaken these factors may be found to be valuable. Likewise, if extensive collecting was a possibility, anatomical evidence is a likely candidate for providing more synapomorphies. Callixylon tracheids exhibit a characteristic patterning of pits in the radial walls that may be a source of variation. Currently the occurrence of anatomy is too patchy to be of use.
It seems to me that it is at least theoretically possible for phylogenetic analysis to be able to distinguish between individuals of the same population if it is provided with sufficient information. However, many arguments have been raised that suggest that inputting data on correlated characters is simply giving more weight to the first character. It is believed by this author though that by taking this to an extreme and minutely describing each specimen to the software a more accurate picture of the variation involved will result. The question does arise at this point however of whether this has become a case of using parsimony to do phenetic analysis.
In a study of fossil taxa there is always a nagging feeling of barking up the wrong tree all together, usually because of the fragmentary nature of the specimens and thus the data. These considerations could be minimised if collections were made that have sample sizes as large as extant population biology studies. It is believed that sufficient material of Archaeopteris exists, based on the collections that have already been made and the cosmopolitan occurrence of Archaeopteris. Assuming that this could be done, at least for one population of Archaeopteris, an evaluation of the interspecific and intraspecific variation could be carried out using multivariate analysis that would hopefully help to alleviate the ranking part of this phylogenetic problem.
Before becoming too delirious with ideas and fantasies it must be acknowledged that many assumptions have been made. The logistic problems of a study like this and the need for extensive collecting are clearly understood. However, the goal of this study was to see if it is feasible to carry out phylogenetic analysis on fossil taxa. With this in mind the above recommendations outline a procedure that will at least summarize the variation of fossil taxa as objectively as possible.
The information generated from such a study, if the study were really inclusive of all known locations of Archaeopteris, would be of potentially great benefit to biogeographers and paleoecologists. Good variation and distribution data may reveal more about the mid-late Paleozoic climate patterns, and the extent of cover of the continents by Archaeopteris forests may become more clear.
References Cited
Arnold, Chester A. (1939). Observations, etc. IV. Plant remains from the Catskill Delta deposits of northern Pennsylvania and southern New York. Contrib. Mus. Paleon. Univ. Mich. 5: 271-313.
Beck, Charles B. (1960). Connection between Archaeopteris and Callixylon. Science 131: 124-125.
Beck, Charles B. (1961 ). The identity of Archaeopteris and Callixylon. Brittonia 12: 351-68.
Dawson, Sir J. W. (1871 ). The fossil plants of the Devonian and Upper Silurian Formations of Canada. Quart. Journ. Geel. Survey Canada, p.1-92., Montreal.
Kenrick, Paul and Muriel Fairon-Demaret (1991). Archaeopteris roemeriana (Geppert) sensu Stockmans, 1948 from the Upper Famennian of Belgium: anatomy and leaf polymorphism. Bulletin de l’lnstitut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 61 :179-195.
Krausel, R. and H. Weyland (1941). Pflanzenreste aus dem Devon von Nord Amerika. Palaeontographica 86B: 1-78.
Mishler, Brent D. and Robert N. Brandon (1987). Individuality pluralism and the phylogenetic species concept. Biology and Philosophy 2: 397-414.
Stewart, Wilson N. (1983). Paleobotany and the evolution of plants. Cambridge University Press.
Zimmerman, W. (1952). The main results of the “telome theory.” The Palaeobotanist, 1: 456·470.
